")
")
مترجم: حبیب الله علیخانی
منبع:راسخون
منبع:راسخون
ویروس های توموری DNA و RNA که در دهه ی 1970 کشف شدند، موجب شده اند تا تئوری ساده و قدرتمندی در مورد تومورهای انسانی ایجاد شود. ویروس هایی که به صورت متداول در بدن انسان ها ایجاد می شوند، بافت های مستعد را آلوده می کند و موجب می شود تا سلول های آلوده، تغییر شکل دهند. این سلول ها در حقیقت، شروع به تکثیر می کنند و دیر یا زود، توده های سلولی تشکیل می دهند که در کلینیک انکولوژی قابل تشخیص می باشند. از آنجایی که سلول های توموری تنها با ورود ژن های اندک، موجب ایجاد تغییر در سلول های جوندگان و مرغ و ایجاد سلول های توموری می شود، این سلول های توموری قابلیت ایجاد تغییر در سلول های انسانی را نیز دارا می باشند.
با گذر زمان، این سناریو مورد توجه قرار گرفت زیرا زیست شناسی و اپیدمیولوژی مربوط به سرطان های انسانی، به سختی منطبق می شوند. بیشتر انواع سرطان های انسانی، به طور رواضح از یک فرد به فرد دیگر انتقال نمی یابند (برخلاف بیماری های عفونی). موارد اندکی از شیوع بیماری های سرطانی، گزارش شده است. حتی از این مهم تر، تلاش های انجام شده در طی دهه ی 1970 به منظور ایزوله کردن ویروس ها از بیشتر انواع تومورهای انسانی، موفقیت آمیز نبوده است. از بین صد ها و یا انواع بیشتر از تومورهای مشاهده شده در کلینیک انکولوژی، تنها دو نوع از آنها که در جوامع غربی شیوع دارند، به طور واضح مربوط به عوامل ویروسی خاص هستند.
این واقعیت موجب ایجاد دو پاسخ می شود. کسانی که به ویروس های توموری به عنوان عوامل مسبب تمام سرطان های انسانی، متوسل می شوند، استدلال کرده اند که عوامل شیمیایی و فیزیکی با ویروس هایی واکنش می دهند که به طور نرمال در داخل سلول های بدن، وجود دارد و موجب فعال شدن قدرت سرطان زایی آنها می شود. سایر محققین پاسخ خود را با کنار گذاشتن کامل ویروس ها، ارائه کرده اند و به دنبال منابع دیگری از ژن ها گشتند که موجب بروز سلول های سرطانی می شوند. آنها این ژن ها را در میان ژنوم سلولی دارای هزاران ژن، دنبال می کنند. این مورد آخر موفقیت آمیز بود و در انتهای دهه ی 1980، ژنوم سلولی به عنوان یک منبع غنی از ژن هایی تشخیص دادند که از تکثیر سلول های سرطانی انسانی ایجاد می شوند.
بنابراین، ویروس های توموری، وقتی به عنوان عوامل کلیدی ایجاد کننده ی سرطان انسانی، در نظر گرفته شوند، نمی توانند انتظارات مربوطه را برطرف کند. از قضا، تحقیقات بر روی ویروس های توموری، توانست کمک زیادی به کشف ژن های سلولی می کند که مسئول فنوتیپ های سلولی نئوپلاستی هستند. گروه قابل توجهی از ژن های عامل سرطان، در طی دهه های متمادی کشف شدند. این ژن ها، آنکوژن ها و ژن های سرکوب کننده ی تومور هستند که به طور مستقیم در تلاش های انجام شده به منظور بررسی عوامل سرطان زا در جمعیت های انسانی، کشف شده اند.
آیا سرطان بوسیله ی فعال سازی رتروویروس درون زا شروع می شود؟
تحقیقات انجام شده در ژاپن بوسیله ی Katsusaburo Yamagiwa که در دهه ی ابتدای قرن 20 ام شروع شده است، نشان داده است که عوامل شیمیایی می توانند موجب القای سرطات در حیوان های آزمایشگاهی شود. همانگونه که قبلا گفته شد، کار او نشاندهنده ی این است که بعد از چند ماه قرارگیری قطران زغال سنگ بر روی گوش های موش ، سرطان پوست در این موش ها ظاهر می شود. در میانه ی این دهه، یک تز دکترا در پاریس در مورد 100 مورد سرطان انسانی، ارائه شده است که بیشتر این سرطان ها، سرطان پوست بوده است. سرطان پوست به دلیل در تماس بودن این افراد با اشعه ی ایکس، ایجاد شده است. بنابراین، واضح است که این عوامل، به طور مستقیم موجب بروز تومورهایی شده است که بیولوژیک نبوده است و در واقع بواسطه ی عوامل شیمیایی یا تابش، حاصل شده است.
این کشف ها در میانه ی قرن 20 ام، برای همه ی محققین سرطان واضح شده بود و آنها، این تئوری را قبول کرده بودند که تمام سرطان ها، از یک طریق ایجاد می شود و یا بوسیله ی عمل مربوط به عوامل عفونی ایجاد می شود که این عوامل، ویروس های توموری هستند. در پاسخ به این مورد، برخی از پیروان نظریه ویروس سرطانی، مخصوصاً آنهایی که بر روی رتروویروس ها کار می کردند، یک مکانیزم جدید در اوایل دهه ی 1970 پیشنهاد کردند. مدل آنها توضیح می دهد که چگونه ویروس های توموری می توانند در تشکیل بسیاری از سرطان هایی مشارکت کننند که علائم ظاهری مربوط به عفونت ویروسی را ندارند.
این روش جدید از زیست شناسی عجیب و غریب رتروویروس ها، مشتق شده است. به خاطر همین مسئله، ژنوم رتروویروس ها در کروموزوم های خط ژرم مربوط به گونه های مختلف مهره داران تجمع می یابند و پرو ویروس های منتج شده (proviruses) سپس مشابه با آلل مندلی، از نسلی به نسل دیگر، انتقال می یابد. در اغلب موارد، این پروویروس های درون زا در گذر از نسل ها، مخفی هستند و حضور آنها در تمام سلول های مربوط به یک ارگان، ظاهر نمی شود. به هر حال، در برخی موارد، این ممکن است که بیان این پروویروس ها درون زای مخفی، ظاهر شود. این مسئله اغلب موجب حفظ قابلیت کدگذاری ذرات ویروس های عفونی می شود.
فعال سازی یک ژنوم ویروسی درون زا (ERV) در فیبروبلاسم بدست آمده از برخی از گونه های خاص موش آزمایشگاهی، می تواند با کشت این سلول ها در حضور تی میدین (thymidine) و برموداکسیاوریدین (BrdU) (bromodeoxyuridine)، ایجاد شود. در پاسخ، این سلول های بافت پیوندی، که بظاهر عاری از عفونت ویروسی است، به طور ناگهانی، شروع به آزاد سازی ذرات رتروویروس می کند که علت آن، سرکوبی مجددی تکثیر مربوط به حالت نهان آنهاست. به طور مشابه، پروویروس های درون زای نهان، ممکن است به طور ناگهانی و در داخل بدن، فعال سازی شوند. این مسئله در تعدلد اندکی از سلول های یک موش، انجام می شود. وقتی ذرات ویروس عفونی، از این سلول های اندک آزاد می شوند، آنها می توانند عفونت سلول به سلول، ایجاد کنند و به سرعت در میان بدن گسترش یابد. این مسئله موجب بروز سرطان در این حیوان ها می شود.
دانستن رفتار رتروویروس های درون زا، برخی پیشنهاد کرده اند که سرطان های انسانی، ممکن است در روشی مشابه ایجاد شود. برای مثال، مواد سرطان زا جهش زا مانند آنهایی که در قطران تنباکو وجود دارد، ممکن است موجب تحریک فعال سازی رتروویروس های درون زای نهفته شود. ذرات ویروسی منتج شده، سپس تکثیر می شوند و در تمام بدن فرد منتشر می شوند و مشابه با رترویروس های درون زا در برخی از گونه های موش، این مسئله موجب بروز سرطان در یک یا چند بافت مستعد می شود. در زمان یکسان، در حالی که قابلیت به انتشار در میان بافت های بدن، موجب می شود تا یک چنین ویروس هایی ممکن است قادر به انتشار افقی به افراد دیگر می شود. این مورد این مسئله را توضیح می دهد که چرا سرطان مشابه با بیماری مسری عمل نمی کند. یکی دیگر از مباحث مطرح شده، این است که رتروویروس ها، آنکوژن های ویروسی در خطوط ژرم گونه های مختلف، ایجاد می کنند و این آنکوژن های ویروسی نهان، با انواع مختلف از مواد سرطان زا، فعال می شوند.
در عین جالب بودن، این مدل های علت شناسی مربوط به سرطان های انسانی، به زودی دچار سقوط و اضمحلال شدند زیرا شواهد تأیید کننده در مورد آنها وجود ندارد. گزارش ها در مورد ذرات ویروسی عفونی در تومورهای انسانی نمی توانند مورد تأیید قرار گیرند. حتی ذرات ویروسی حاوی ترانس کریپتاز معکوس نیز به سختی در تومورهای انسانی یافت می شوند.
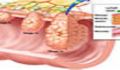
این واضح شده است که بیشتر ژنوم ویروسی درون زا که در ژنوم انسانی وجود دارند، اثری از عفونت های خط جرام است که در حدود 5 میلیون سال پیش در پستانداران آبا و اجدادی ما بوجود آمده است. از آن زمان، این پیروویروس ها، در توالی ها ایجاد جهش کرده اند و دیگر قادر به تشخیص ذرات رتروویروس های عفونی وجود نداشت و بدین وسیله، این بخش ها به قسمت هایی از DNA متصل شده اند که موجب تشکیل توده ی ژنوم ماده شده است (شکل 1). اگر چه 8 % از ژنوم های انسانی، از ژنوم ویروسی درون زا نشئت می گیرند، تنها چند تا از 40000 بخش مشتق شده از رتروویروس ها، قادر است تا ذرات ویروسی عفونی ایجاد کند. یکی از زیرشاخه های این ویروس ها، که خانواده ی HERV-K نامیده می شوند، اخیراً وارد خط ژنومی انسانی شده اند و چند تا از پیروویروس های آنها ظاهراً سالم هستند اما امروزه، حتی این نوع از بخش ها نیز ویروس های عفونی تولید نمی کنند و در سلول های سرطانی متحرک شده اند. بنابراین، محققین سرطان به جایی چشم دوختند که المان های ژنتیکی ممکن است موجب تشکیل سرطان شوند.
") ترانس فکشن DNA یک استراتژی برای تشخیص آنکوژن های غیر ویروسی مهیا می کند
ترانس فکشن DNA یک استراتژی برای تشخیص آنکوژن های غیر ویروسی مهیا می کند
برای این محققین که قصد دارند تا در مورد سرطان غیر ویروسی اطلاعات کسب کنند، رد نظریه رتروویروس درون زا موجب می شود تا یک نظریه قابل دوام بر جای بماند. با توجه به این تئوری، عملکرد آنکوژنی به عنوان عوامل جهش زا در نظر گرفته می شود. خواه این عوامل فیزیکی باشند و یا خواه شیمیایی، این عوامل موجب القای سرطان از طریق قابلیت جهش زایی ژن های کنترل کننده ی رشد در ژنوم های مربوط به سلول های مستعد، می شود. وقتی این ژن ها جهش زا شوند، آلل های جهش زای منتج شده، ممکن است به عنوان آنکوژن های فعال عمل کنند و بدین وسیله، رشد سرطانی سلول ها رخ می دهد.
این مدل به صورت متفاوت پیش بینی می کند که سلول های تغییر شکل یافته ی شیمیایی، ژن های جهش یافته ای حمل می کنند و این ژن ها، مسئول برنامه ریزی رشد غیر نرمال این سلول ها، هستند. در حقیقت، این مفهوم که سلول سرطانی، سلول های جهش یافته ای هستند، در شروع قرن 20 ام مطرح شد و بوسیله ی David von Hansemann و Theodor Boveri مطرح گردیده است. این ممکن است تا تعداد یک چنین ژن های جهش یافته را در ژنوم این سلول ها، پیش بینی نمود. از این مهم تر، اثبات های تجربی در مورد وجود ژن های ایجاد کننده ی سرطان به اثبات رسید. این مسئله یک چالش دلهره آور ایجاد می کند. اگر این ژن ها واقعا در ژنوم سلول هایی وجود داشته باشد که از لحاظ شیمیایی تغییر شکل یافته اند، چگونه این ژن ها تشکیل می شوند؟ اگر این ژن ها ورژن جهش یافته ای از ژن های سلولی نرمال باشند، بنابراین، آنها در ژنوم سلول های سرطانی به همراه هزاران و یا صدها هزار ژن دیگر، وجود دارند (که در هر ژنوم هاپلوئیدی، یک کپی وجود دارد). این ژن های سرطانی، در صورت موجود، به طور واضح سوزنی شکل هستند.
برای تعیین این مسئله که آیا انکوژنهای غیر ویروسی در سلول های تغییر شکل یافته ی شیمیایی وجود دارد، استراتژی های تجربی جدیدی پیشنهاد شده است. این مورد شامل ورود DNA ی مربوط به سلول های سرطانی به داخل سلول های نرمال است و سپس تعیین می کند که آیا سلول های گیرنده در پاسخ به ورود DNA ی سلول توموری، تغییر شکل می دهند یا نه؟ یک چنین تغییری می تواند بوسیله ی ظاهر کانون در محیط های کشت سلول های گیرنده، تشخیص داده شود. این استراتژی به چندین مزیت تجربی، وابسته است که شامل توسعه ی یک رویه ی انتقال ژن مؤثر، می باشد. علاوه بر این، استخراج DNA از سلول های سرطانی مناسب و سلول های گیرنده ی مطلوب، برای تشخیص، ضروری می باشد.
در سال 1972، یک رویه ی انتقال مؤثر ژن ابداع شد که بعدها، به آن ترانس فکشن (transfection) گفتند. این رویه که بوسیله ی آن قابلیت ورود مولکول های منفرد DNA به داخل سلول های پستانداران وجود دارد (شکل 1)، بر اساس هم رسوبی DNA با کریستال های کلسیم فسفات، کار می کند. به دلایلی که هم اکنون نیز به خوبی شناخته نشده است، این هم رسوبی می تواند بر روی مقاومت ذاتی سلول ها در برابر مواد ژنتیکی خارجی، فایق آید. این مشخص شده است که سلول های موجود در خط سلول NIH 3T3 که در اصل از فیبروبلاست جنین موش بدست آمده اند، به عنوان DNA خارجی وارد شده به این بخش ها، در نظر گرفته شده است.
این موضوع آخر پیش از این مسئله حل و فصل شد که برنامه ی تجربی بتواند سلول های سرطانی دهنده ای را تشخیص دهد که از آن DNA آماده سازی می شود. در اینجا، محققین کورکورانه عمل کرده اند. این واضح نیست که آیا تمام انواع سلول های سرطانی دارای ژن های تغییر شکل یافته مانند آنکوژن src هستند یا نه؟ همچنین این فهمیده نشده است که آیا یک ژن تغییر شکل یافته ی سلولی (یک آنکوژن سلولی) که تبدیل سلول های اپیتلیال طبیعی به سلول های سرطان است، همچنین قادر به این است که عملکردی غیر مشابه در محیط های بین سلولی مربوط به بافت اتصالی داشته باشد یا نه؟ البته هنوز هم مسائل ممکنه ی دیگری وجود دارد. برای مثال، یک آنکوژن که مسئول تغییر شکل سلول های انسانی نرمال به سلول های سرطانی است ممکن است نتواند سلول های نرمال موش را تغییر شکل دهد، علت این مسئله برخی از ناسازگاری های خاص است.
با در نظر گرفتن این مسائل در ذهن، محققین سلول های توموری دهنده ای را در نظر گرفته اند که از فیبروبلاست موش مشتق شده اند. این سلول های سرطانی خاص از فیبروبلاست موش و از خط سلولی C3H10T1/2 موش مشتق شده اند که به صورت تکراری با استفاده از عوامل سرطان زا و تری متیل کلانترن (3-MC) عمل آوری شده اند. تری متیل کلانترن یک ترکیب شناخته شده از قطران ذغال سنگ است. از این مهم تر، این سلول ها از بخش هایی مشتق شده اند که در آن هیچ عفونت ویروسی و یا ژنوم ویروسی درون زا مشاهده نشده است. در اینجا، هر آنکوژن تغییر شکل دهنده ی تشخیص داده شده در ژنوم این سلول ها، منشأ داخلی دارند. این منشأ داخلی، در واقع نسخه جهش یافته از ژن سلولی نرمال هستند.
در بین سال های 1978- 1979، DNA های مشتق شده از چند خط سلولی تغییر شکل یافته بوسیله ی 3-MC، به داخل محیط کشت های محتوی سلول های گیرنده ی NIH 3T3 انتقال یافت. این مسئله منجر به تولید یک تعداد قابل توجه از کانون های تمرکز سلولی در طی چند هفته می شود. سلول های بدست آمده از بخش متمرکز بعدها موجب ایجاد تومور می شوند (جدول 1). این آزمایش ساده، اثبات کرد که DNA تومور دهنده که حامل یک یا چند المان ژنتیکی است، قادر است تا به صورت سلول گیرنده ی NIH 3T3 غیر توموری تبدیل شود که این سلول ها، به شدت تومورزا هستند.
DNA استخراج شده از سلول های نرمال و غیر تغییر شکل یافته ی C3H10T1/2 قادر به القای حالت های کانونی در مونولایه های سلولی NIH 3T3 نیستند. این تفاوت موجب می شود تا سلول های نرمال C3H10T1/2 که قبلا تحت عامل سرطان زای 3-MC هستند، ژنومی با ساختار متفاوت داشته باشند. این مسئله منجر به ایجاد توالی های ژنی جدیدی می شود که دارای قدرت تغییر شکل هستند. به عبارت دیگر، این محتمل به نظر می رسد که عامل سرطان زای 3-MC یک ژن یا ژن های نرمال با نام C3H10T1/2 را به آلل های جهش زا تبدیل می کند. این آلل ها وقتی وارد سلول های NIH3T3 می شوند، نقش آنکوژن های تغییر شکل دهنده را بازی می کنند.
در ابتدا، این به نظر می رسد که تعیین این مسئله سخت است که آیا سلول های توموری دهنده، حامل یک آنکوژن منفرد در ژنوم خود هستند یا چندین آنکوژن متمایز وجود دارد که به صورت هماهنگ عمل کرده و موجب تغییر شمل در سلول های گیرنده می شوند. آنالیز دقیق انجام شده بر روی ترانس فکشن این مسئله را حل کرد. محققین کشف کردند که وقتی DNA سلولی به سلول گیرنده وارد شود، تنها حدود 0.1 % از ارزش ژنوم سلولی مربوط به DNA ی دهنده، وارد ژنوم هر یک از سلول های گیرنده ی تغییر شکل یافته می شود. احتمال اینکه دو ژن دهنده ی غیر مشابه و مستقل از لحاظ ژنتیکی، وارد یک سلول گیرنده ی منفرد شوند، برابر با 〖10〗^3×〖10〗^3 می شود. این احتمال بسیار بعید است. با استفاده از این محاسبات، آنها می توانند استدلال کنند که تنها یک ژن منفرد مسئول تغییر شکل سلول های NIH 3T3 (بعد از ترانس فکشن DNA سلول توموری دهنده) است. در حقیقت، این مسئله منجر این استدلال می شود که سال ها پیش از اینکه برخورد سلول های نرمال C3H10T1/2 موش با عامل سرطان زای 3-MC، موجب تشکیل آلل آنکوژن جهش زای منفرد شود، این آلل ها قادر به تغییر شکل در هر دو سلول C3H10T1/2 و سلول های گیرنده ی NIH 3T3می شود که در این حالت، این آلل ها بوسیله ی انتقال دهنده های ژنی ایجاد می شوند.
این تجربیات مربوط به ترانس فکشن اهمیت بالایی داشته است و بوسیله ی آن، شاخصی مناسب برای مسئله ی آنکوژن ها، مطرح شده است. همچنین این فهمیده شده است که هیچ ارتباط ظاهری میان مکانیزم های مربوط به ژنوم سلولی و عفونت ویروسی وجود ندارد. شاید سلول های تومور انسانی که احتمالا از طریق مکانیزم های غیر ویروسی ظاهری می شوند، همچنین آنکوژن های با قابلیت ترانس فکشن، ایجاد می کنند. آیا آنکوژن های حیوانی (در صورتی که در ژنوم این سلول ها وجود داشته باشد)، همچنین قادر به تغییر رفتار سلول های موش هستند یا نه؟
این سوال و سوالات مشابه به زودی پاسخ داده شد. جواب این سوال ها و سوالات مشابه، جواب مثبت می باشد. DNA های استخراج شده از خط سلولی سرطان های مثانه، ریه و روده یک انسان و DNA مربوط به لوسمی همگی قادر به تغییر شکل در فیبروبلاست NIH3T3 موش گیرنده، هستند (شکل 2). این بدین معناست که آنکوژن های موجود در خطوط سلولی (با هر طبیعتی که دارند)، قادر به تغییر شکل سلولی هستند.
آنکوژن های کشف شده در خطوط سلولی تومور انسانی با آنکوژن هایی در ارتباط است که بوسیله ی رتروویروس های تغییر شکل دهنده، حمل می شوند
آنکوژن های تشخیص داده شده بوسیله ی ترانس فکشن در ژنوم های مختلف سلول های تومور انسانی ظاهرا از ژن های سلولی نرمال قبلی مشتق شده اند که در آنها عملکرد آنکوژن درست نیست. این به نظر می رسد که به موازات این فرایند که منجر به ایجاد رتروویروس های ایجاد کننده ی تغییر شکل، در طی تشکیل این ویروس ها، ژن های سلولی نرمال یعنی پروتو- آنکوژن ها فعال می شوند، ولو اینکه یک مکانیزم ژنتیکی متفاوت رخ دهد.
این توازی ظاهری منجر به مطرح شدن یک سوال واضح می شود: آیا گروه مشابه از پروتو- آنکوژن های سلولی بواسطه ی تلاش رتروویروس ها و سایر عوامل جهش زای غیر ویروسی، فعال سازی می شود؟ یا آیا آنکوژن های مربوط به رتروویروس ها و آنهایی که بوسیله ی مکانیزم های فعال سازی، ایجاد شده اند، از دو گروه متمایز از پروتو- آنکوژن ها می باشند؟
استفاده از پروب های DNA مختص آنکوژن های رتروویروسی، جواب این پاسخ را ارائه می دهد. با استفاده از روش blot جنوبی (Southern blot procedure)، یک پروب DNA که از آنکوژه های H-ras مشتق شده است، قادر به تشخیص و تشکیل ترکیبی از آنکوژن ها می باشد. این آنکوژن ها در DNA مربوط به سلول لوسمی مثانه ی یک انسان، بدست آمده اند (شکل 3). یک آنکوژن مربوطه، که آنکوژن K-ras نامیده می شود و در ژنوم ویروس تومور بدخیم بافت هم بند وجود دارد، قادر است تا با آنکوژن تشخیص داده شده بوسیله ی ترانس فکشن DNA (از یک خط سلولی سرطان روده ی انسانی) تحت تابش قرار گیرد.
لیست مربوط به ارتباط میان آنکوژن های مربوط به رتروویروس و آنکوژن های موجود در تومورهای ایجاد شده ی غیر ویروسی، به زودی رشد داده شد (جدول 2). در این موارد، ارتباط های معمولا بعد از کشف این مسئله به فراموشی سپرده شد که آنکوژن های مربوط به رتروویروس ها، در برخی کپی های مربوط به ژنوم سلول های سرطان انسانی، وجود دارد. آنکوژن myc که در اصل در کپی های مضاعف ژن myc مشاهده شده است، منجر به ایجاد فرایند تقویت ژنی (gene amplification) می شود و ظاهراً موجب افزایش سطح محصول پروتئینی آنکوژن myc می شود. این پروتئین اضافی موجب افزایش تکثیر سلول های سرطانی می شود. این فهمیده شده است که ژن erbB که اولین بار بوسیله ی وجود آن در ژنوم ویروس ارویتززوبلاستوتیز مرغی (avian erythroblastosis virus) شناخته شده است، موجب افزایش تعداد کپی های موجود در DNA ی مربوط به سلول های توموری شکم، سینه و مغز می شود. میزان بالای ژن مشابه با erbB هم اکنون در بیشتر سرطان ها، گزارش شده است.
در سال 1987، تقویت ژن های مربوط به erbB که هم اکنون آن را erbB2 ، HER2 و یا neu نامیده می شود، در بسیاری از سرطان های سینه، گزارش شده است (شکل 4A). افزایش در تعداد کپی های ژنی به میزان بیش از 5 کپی در هر سلول سرطانی مشاهده می شود. این مسئله با کاهش در تعداد بیماران، مرتبط است. در واقع بعد از درمان اولیه، دیگر نیازی به درمان دیگری نداشتند (شکل 4B). به طور قابل توجه، تقویت مشاهده شده در ژن erbB2/HER2 اغلب و نه همیشه، در ارتباط با افزایش میزان پروتئین های کدگذاری شده می شود. در میان یک گروه بزرگ از بیماران مبتلا به سرطان سینه، آنهایی که دارای تومورهایی هستند که در آنها سطح این پروتئین بالاست، 6 تا 7 سال بعد از تشخیص زنده بوده اند، در حالی که بیمارانی که دارای تومورهایی هستند که در آنها سطح بالایی از این پروتئین وجود دارد، عمری 3 ساله بعد از تشخیص داشته اند. این ارتباط معکوس میان سطح erbB2/HER2 و زنده ماندن طولانی بیماران، شاخصی مهم در مورد این مسئله است که این ژن در حالت تقویت شده، موجب رشد بدخیم سلول های سرطانی می شود.
در سال های بعدی، با استفاده از روش های جدیدی مانند روش هیبریداسیون فلورسانس در جا (in situ hybridization (FISH) برای تعیین میزان تقویت ژنی در این نوع و انواع دیگر سلول ها، استفاده شد. در سال 2010، یک سری روش های دیگر برای بررسی و تأیید اثر آنکوژن ها بر روی تقویت ژنی در سرطان های انسانی، مورد استفاده قرار گرفت (شکل 4C). این کار منجر به رسیدن به لیستی 77 تایی از ژن ها شد که به احتمال زیاد، در ایجاد تومور اثر دارند.
همین طور که روش های ژنتیکی پیشرفت کردند، قابلیت بررسی کل ژنوم سلولی تومور در نواحی کروموزومی که از این مشکلات رنج می بردند، به صورت قابل توجهی، بهبود یافت و بررسی های گسترده ای بر روی تقویت و حذف ژنی انجام شد. امروزه، این روش ها، به صورت روش های متداول در آمده است. در مورد سلول های سرطانی، این تقویت ها، منجر به افزایش میزان ژن های افزایش دهنده ی رشد (پروتو-آنکوژن ها) می شود، در حالی که حذف این ژن ها، موجب کاهش ژن های جلوگیری کننده در برابر رشد می شود. همانگونه که در شکل 5 نشان داده شده است، در طی تغییر شکل، انواع مختلف سلول ها متحمل تغییرات ژنتیکی مختلفی می شوند.
آلل های جهش زایی که از آنکوژن src مشتق شده است، اولین آنکوژن سلولی بوده است که کشف شده است. اثبات شده است که این آلل ها از ژنوم سلولی مربوط به تومورهای انسانی گریزان هستند. در نهایت، در سال 1999، تقریبا یک ربع قرن بعد از تکثیر ژن src، شکل جهش زای ژن src در ژنوم سلول های تومور انسانی، کشف شد. این شکل مخصوصا در ژنوم 12 % از سرطان های پیشرفته ی روده ی انسانی، مشاهده می گردد.
یک وضعیت کاملا متفاوت، بعد از این مشاهده شده است که پروتو- آنکوژن ها بوسیله ی یک رتروویروس کشف شود. بعد از ورود به ژنوم رتروویروس، میزان این ژن دریافت شده، بوسیله ی یک پروموتر رونویسی ویروسی کنترل می شود. این مسئله موجب افزایش میزان ژن می شود. نسخه برداری این ژن مربوط به ویروس، که اکنون به آن آنکوژن گفته می شود، دیگر پاسخی برای سیگنال های سلولی نیست که قبلا میزان آن را تنظیم می کرد. برای مثال، در مورد c-myc میزان این ژن به طور نرمال، بوسیله ی تغییر در سطح سیگنال های خارج سلولی، کنترل می شود مانند چیزی که بوسیله ی فاکتورهای رشد میتوژنی هدایت می شود. وقتی این مورد در ژنوم ویروس میلوسیتوماتوزیز مرغی (avian myelocytomatosis virus (AMV)) مشاهده شود، میزان آن نسبت به حالت نرمال در این سلول ها، بالاست و این میزان در سطح ثابتی قرار دارد.
اما چطور پروتو- آنکوژن H-ras به آنکوژن مناسب تبدیل شود که بوسیله ی ترانس فکشن DNA ی سرطان مثانه ی انسانی، قابل تشخیص باشد؟ تقویت ژنی نمی تواند فعال سازی آن را تضمین کند زیرا این به نظر می رسد که این آنکوژن در DNA سرطان مثانه به عنوان یک ژن کپی شده ی منفرد، وجود دارند. این پازل در زمانی که این آنکوژن سرطان مثانه ی H-ras بوسیله ی کلونی های مولکولی ایزوله شدند، رشد می کند. این آنکوژن در یک بخش DNA ژنومی متمرکز می شوند که دارای طول 6.6 کیلوباز (kilobases) است. این مسئله جالب توجه است که یک بخش DNA با اندازه ی ایده آل، در DNA های انسانی نرمال، مشاهده می شود. بخش آخر به طور واضح بیان کننده ی پروتو- آنکوژن های H-ras می باشد. این آنکوژن یک ژن نرمال است که از برخی جهش ها رنج می برد و در طی تشکیل سرطان مثانه، به آنکوژن تبدیل شده است.
در حالی که ساختارهای کلی DNA بسیار مشابه است، این دو نوع مختلف از ژن H-ras به شیوه ی متفاوتی عمل می کنند. آنکوژنی که از سلول های سرطان مثانه ی انسانی بدست آمده است، موجب تغییر در سلول های NIH 3T3 می شود، در حالی که پروتو- آنکوژن نرمال این قابلیت را ندارند. این راز به نقشه ی جزئی مربوط به مکان های کلیواژی وابسته است که مربوط به آنزیم های محدودکننده ی مختلف می باشد. این مسئله نشان داده است دو نوع مختلف از ژن ها، دارای ساختار فیزیکی هستند که قابل تشخیص هستند.
هنوز این واضح است که دو نوع مختلف از ژن های H-ras دارای برخی تفاوت های قابل توجه در توالی خود هستند. علت این مسئله، عامل های مختلف آنهاست. تفاوت های موجود در توالی بحرانی آنها در اصل بوسیله ی ترکیب مجدد بخش های پروتو- آنکوژن با سایر بخش های مشتق شده از آنکوژن، محدود می شود (شکل 6). این مسئله موجب می شود تا تفاوت حیاتی میان یک بخش، تنها 350 جفت باشد.
این پازل در نهایت در زمانی حل شد که بخش های 350-bp مربوطه از پروتو-آنکوژن و آنکوژن مورد آنالیز توالی DNA قرار گرفتند. تفاوت اساسی در این دو مورد، فوق العاده ظریف است. این تفاوت در واقع یک جانشینی اساسی منفرد است که در آن، یک G (گوانوزین) در آنکوژن با یک T (تیمیدین) جایگزین می شود. این جایگزینی منفرد (یک جهش نقطه ای)، موجب می شود تا آنکوژن به پروتوآنکوژن تبدیل شود (شکل 7). این کشف مهم در سه آزمایشگاه انجام شد و تمام شک ها در مورد درستی آن را از بین برد.
کشف این جهش نقطه ای، نقطه ی تعیین کننده ای در تحقیقات سرطان می باشد. این اولین زمان بود که یک جهش در یک ژن کشف شد که موجب رشد نئوپلاستیک سرطان در انسان می شود. این مسئله نیز مهم است که این تغییر ژنتیکی به عنوان یک جهش سوماتیک (somatic mutation) در نظر گرفته می شود.
با توجه به این اطلاعات، محققین می توانند یک توضیح محتمل در مورد منشأ سرطان مثانه ارائه دهند و با تعمیم این نظریه، می توان این توضیح را در مورد منشأ تمام سرطان ها، ارائه کرد. سرطان های خاص مانند سرطان مثانه که در آن آنکوژن H-ras حالت کلونی دارد، در مردان میان سال مشاهده می شود. این افراد در حدود 4 دهه سیگار کشیده اند. در طی این زمان، مواد سرطان زای موجود در سیگار وارد ریه های آنها شده است و با عبور از این محل و ورود به سیستم خون، وارد کلیه های این افراد شده است. این مواد از طریق اوره، دفع می شوند. در حالی که در مثانه، برخی از مولکول های سرطانی، که در داخل اوره قرار دارند، وارد سلول های دیواره ی مثانه می شوند و بر روی DNA این سلول ها، جذب می شوند. در یک زمان، این مواد سرطان زا می توانند موجب بروز یک جهش نقطه ای در پروتوآنکوژن H-ras مربوط به سلول اپیتلیال می شود. بعد از آن، این سلول جهش یافته و نسل های بعدی آن، به صورت کنترل نشده تکثیر می کنند و در هر کدام از آنها، این مسئله وجود دارد. سال های بعد، یک توده ی سرطانی در این بیمار، تشخیص داده می شود.
از این مهم تر، این جایگزینی جفتی در چارچوب ژن H-ras رخ می دهد. به طور خاص، این جهش نقطه ای موجب جایگزینی یک باقیمانده ی گلیسین در باقیمانده های والین مربوط به پروتئین های کد گذاری شده ی H-ras می شود.
کشف این جهش نقطه ای، یک مکانیزم در مورد فعالیت انکوژنی ارائه کرده است که کاملا از فعالیت انکوژنهای MYC متفاوت است. در مورد H-ras، یک تغییر در ساختار مربوط به پروتئین کدگذاری شده، یک مورد حیاتی می باشد. برخلاف آنکوژن MYC، تنظیم نبودن میزان این ماده در قدرت
استفاده از مطالب این مقاله، با ذکر منبع راسخون، بلامانع می باشد.
با گذر زمان، این سناریو مورد توجه قرار گرفت زیرا زیست شناسی و اپیدمیولوژی مربوط به سرطان های انسانی، به سختی منطبق می شوند. بیشتر انواع سرطان های انسانی، به طور رواضح از یک فرد به فرد دیگر انتقال نمی یابند (برخلاف بیماری های عفونی). موارد اندکی از شیوع بیماری های سرطانی، گزارش شده است. حتی از این مهم تر، تلاش های انجام شده در طی دهه ی 1970 به منظور ایزوله کردن ویروس ها از بیشتر انواع تومورهای انسانی، موفقیت آمیز نبوده است. از بین صد ها و یا انواع بیشتر از تومورهای مشاهده شده در کلینیک انکولوژی، تنها دو نوع از آنها که در جوامع غربی شیوع دارند، به طور واضح مربوط به عوامل ویروسی خاص هستند.
این واقعیت موجب ایجاد دو پاسخ می شود. کسانی که به ویروس های توموری به عنوان عوامل مسبب تمام سرطان های انسانی، متوسل می شوند، استدلال کرده اند که عوامل شیمیایی و فیزیکی با ویروس هایی واکنش می دهند که به طور نرمال در داخل سلول های بدن، وجود دارد و موجب فعال شدن قدرت سرطان زایی آنها می شود. سایر محققین پاسخ خود را با کنار گذاشتن کامل ویروس ها، ارائه کرده اند و به دنبال منابع دیگری از ژن ها گشتند که موجب بروز سلول های سرطانی می شوند. آنها این ژن ها را در میان ژنوم سلولی دارای هزاران ژن، دنبال می کنند. این مورد آخر موفقیت آمیز بود و در انتهای دهه ی 1980، ژنوم سلولی به عنوان یک منبع غنی از ژن هایی تشخیص دادند که از تکثیر سلول های سرطانی انسانی ایجاد می شوند.
بنابراین، ویروس های توموری، وقتی به عنوان عوامل کلیدی ایجاد کننده ی سرطان انسانی، در نظر گرفته شوند، نمی توانند انتظارات مربوطه را برطرف کند. از قضا، تحقیقات بر روی ویروس های توموری، توانست کمک زیادی به کشف ژن های سلولی می کند که مسئول فنوتیپ های سلولی نئوپلاستی هستند. گروه قابل توجهی از ژن های عامل سرطان، در طی دهه های متمادی کشف شدند. این ژن ها، آنکوژن ها و ژن های سرکوب کننده ی تومور هستند که به طور مستقیم در تلاش های انجام شده به منظور بررسی عوامل سرطان زا در جمعیت های انسانی، کشف شده اند.
آیا سرطان بوسیله ی فعال سازی رتروویروس درون زا شروع می شود؟
تحقیقات انجام شده در ژاپن بوسیله ی Katsusaburo Yamagiwa که در دهه ی ابتدای قرن 20 ام شروع شده است، نشان داده است که عوامل شیمیایی می توانند موجب القای سرطات در حیوان های آزمایشگاهی شود. همانگونه که قبلا گفته شد، کار او نشاندهنده ی این است که بعد از چند ماه قرارگیری قطران زغال سنگ بر روی گوش های موش ، سرطان پوست در این موش ها ظاهر می شود. در میانه ی این دهه، یک تز دکترا در پاریس در مورد 100 مورد سرطان انسانی، ارائه شده است که بیشتر این سرطان ها، سرطان پوست بوده است. سرطان پوست به دلیل در تماس بودن این افراد با اشعه ی ایکس، ایجاد شده است. بنابراین، واضح است که این عوامل، به طور مستقیم موجب بروز تومورهایی شده است که بیولوژیک نبوده است و در واقع بواسطه ی عوامل شیمیایی یا تابش، حاصل شده است.
این کشف ها در میانه ی قرن 20 ام، برای همه ی محققین سرطان واضح شده بود و آنها، این تئوری را قبول کرده بودند که تمام سرطان ها، از یک طریق ایجاد می شود و یا بوسیله ی عمل مربوط به عوامل عفونی ایجاد می شود که این عوامل، ویروس های توموری هستند. در پاسخ به این مورد، برخی از پیروان نظریه ویروس سرطانی، مخصوصاً آنهایی که بر روی رتروویروس ها کار می کردند، یک مکانیزم جدید در اوایل دهه ی 1970 پیشنهاد کردند. مدل آنها توضیح می دهد که چگونه ویروس های توموری می توانند در تشکیل بسیاری از سرطان هایی مشارکت کننند که علائم ظاهری مربوط به عفونت ویروسی را ندارند.
این روش جدید از زیست شناسی عجیب و غریب رتروویروس ها، مشتق شده است. به خاطر همین مسئله، ژنوم رتروویروس ها در کروموزوم های خط ژرم مربوط به گونه های مختلف مهره داران تجمع می یابند و پرو ویروس های منتج شده (proviruses) سپس مشابه با آلل مندلی، از نسلی به نسل دیگر، انتقال می یابد. در اغلب موارد، این پروویروس های درون زا در گذر از نسل ها، مخفی هستند و حضور آنها در تمام سلول های مربوط به یک ارگان، ظاهر نمی شود. به هر حال، در برخی موارد، این ممکن است که بیان این پروویروس ها درون زای مخفی، ظاهر شود. این مسئله اغلب موجب حفظ قابلیت کدگذاری ذرات ویروس های عفونی می شود.
فعال سازی یک ژنوم ویروسی درون زا (ERV) در فیبروبلاسم بدست آمده از برخی از گونه های خاص موش آزمایشگاهی، می تواند با کشت این سلول ها در حضور تی میدین (thymidine) و برموداکسیاوریدین (BrdU) (bromodeoxyuridine)، ایجاد شود. در پاسخ، این سلول های بافت پیوندی، که بظاهر عاری از عفونت ویروسی است، به طور ناگهانی، شروع به آزاد سازی ذرات رتروویروس می کند که علت آن، سرکوبی مجددی تکثیر مربوط به حالت نهان آنهاست. به طور مشابه، پروویروس های درون زای نهان، ممکن است به طور ناگهانی و در داخل بدن، فعال سازی شوند. این مسئله در تعدلد اندکی از سلول های یک موش، انجام می شود. وقتی ذرات ویروس عفونی، از این سلول های اندک آزاد می شوند، آنها می توانند عفونت سلول به سلول، ایجاد کنند و به سرعت در میان بدن گسترش یابد. این مسئله موجب بروز سرطان در این حیوان ها می شود.
دانستن رفتار رتروویروس های درون زا، برخی پیشنهاد کرده اند که سرطان های انسانی، ممکن است در روشی مشابه ایجاد شود. برای مثال، مواد سرطان زا جهش زا مانند آنهایی که در قطران تنباکو وجود دارد، ممکن است موجب تحریک فعال سازی رتروویروس های درون زای نهفته شود. ذرات ویروسی منتج شده، سپس تکثیر می شوند و در تمام بدن فرد منتشر می شوند و مشابه با رترویروس های درون زا در برخی از گونه های موش، این مسئله موجب بروز سرطان در یک یا چند بافت مستعد می شود. در زمان یکسان، در حالی که قابلیت به انتشار در میان بافت های بدن، موجب می شود تا یک چنین ویروس هایی ممکن است قادر به انتشار افقی به افراد دیگر می شود. این مورد این مسئله را توضیح می دهد که چرا سرطان مشابه با بیماری مسری عمل نمی کند. یکی دیگر از مباحث مطرح شده، این است که رتروویروس ها، آنکوژن های ویروسی در خطوط ژرم گونه های مختلف، ایجاد می کنند و این آنکوژن های ویروسی نهان، با انواع مختلف از مواد سرطان زا، فعال می شوند.
در عین جالب بودن، این مدل های علت شناسی مربوط به سرطان های انسانی، به زودی دچار سقوط و اضمحلال شدند زیرا شواهد تأیید کننده در مورد آنها وجود ندارد. گزارش ها در مورد ذرات ویروسی عفونی در تومورهای انسانی نمی توانند مورد تأیید قرار گیرند. حتی ذرات ویروسی حاوی ترانس کریپتاز معکوس نیز به سختی در تومورهای انسانی یافت می شوند.
این واضح شده است که بیشتر ژنوم ویروسی درون زا که در ژنوم انسانی وجود دارند، اثری از عفونت های خط جرام است که در حدود 5 میلیون سال پیش در پستانداران آبا و اجدادی ما بوجود آمده است. از آن زمان، این پیروویروس ها، در توالی ها ایجاد جهش کرده اند و دیگر قادر به تشخیص ذرات رتروویروس های عفونی وجود نداشت و بدین وسیله، این بخش ها به قسمت هایی از DNA متصل شده اند که موجب تشکیل توده ی ژنوم ماده شده است (شکل 1). اگر چه 8 % از ژنوم های انسانی، از ژنوم ویروسی درون زا نشئت می گیرند، تنها چند تا از 40000 بخش مشتق شده از رتروویروس ها، قادر است تا ذرات ویروسی عفونی ایجاد کند. یکی از زیرشاخه های این ویروس ها، که خانواده ی HERV-K نامیده می شوند، اخیراً وارد خط ژنومی انسانی شده اند و چند تا از پیروویروس های آنها ظاهراً سالم هستند اما امروزه، حتی این نوع از بخش ها نیز ویروس های عفونی تولید نمی کنند و در سلول های سرطانی متحرک شده اند. بنابراین، محققین سرطان به جایی چشم دوختند که المان های ژنتیکی ممکن است موجب تشکیل سرطان شوند.
")
برای این محققین که قصد دارند تا در مورد سرطان غیر ویروسی اطلاعات کسب کنند، رد نظریه رتروویروس درون زا موجب می شود تا یک نظریه قابل دوام بر جای بماند. با توجه به این تئوری، عملکرد آنکوژنی به عنوان عوامل جهش زا در نظر گرفته می شود. خواه این عوامل فیزیکی باشند و یا خواه شیمیایی، این عوامل موجب القای سرطان از طریق قابلیت جهش زایی ژن های کنترل کننده ی رشد در ژنوم های مربوط به سلول های مستعد، می شود. وقتی این ژن ها جهش زا شوند، آلل های جهش زای منتج شده، ممکن است به عنوان آنکوژن های فعال عمل کنند و بدین وسیله، رشد سرطانی سلول ها رخ می دهد.
این مدل به صورت متفاوت پیش بینی می کند که سلول های تغییر شکل یافته ی شیمیایی، ژن های جهش یافته ای حمل می کنند و این ژن ها، مسئول برنامه ریزی رشد غیر نرمال این سلول ها، هستند. در حقیقت، این مفهوم که سلول سرطانی، سلول های جهش یافته ای هستند، در شروع قرن 20 ام مطرح شد و بوسیله ی David von Hansemann و Theodor Boveri مطرح گردیده است. این ممکن است تا تعداد یک چنین ژن های جهش یافته را در ژنوم این سلول ها، پیش بینی نمود. از این مهم تر، اثبات های تجربی در مورد وجود ژن های ایجاد کننده ی سرطان به اثبات رسید. این مسئله یک چالش دلهره آور ایجاد می کند. اگر این ژن ها واقعا در ژنوم سلول هایی وجود داشته باشد که از لحاظ شیمیایی تغییر شکل یافته اند، چگونه این ژن ها تشکیل می شوند؟ اگر این ژن ها ورژن جهش یافته ای از ژن های سلولی نرمال باشند، بنابراین، آنها در ژنوم سلول های سرطانی به همراه هزاران و یا صدها هزار ژن دیگر، وجود دارند (که در هر ژنوم هاپلوئیدی، یک کپی وجود دارد). این ژن های سرطانی، در صورت موجود، به طور واضح سوزنی شکل هستند.
برای تعیین این مسئله که آیا انکوژنهای غیر ویروسی در سلول های تغییر شکل یافته ی شیمیایی وجود دارد، استراتژی های تجربی جدیدی پیشنهاد شده است. این مورد شامل ورود DNA ی مربوط به سلول های سرطانی به داخل سلول های نرمال است و سپس تعیین می کند که آیا سلول های گیرنده در پاسخ به ورود DNA ی سلول توموری، تغییر شکل می دهند یا نه؟ یک چنین تغییری می تواند بوسیله ی ظاهر کانون در محیط های کشت سلول های گیرنده، تشخیص داده شود. این استراتژی به چندین مزیت تجربی، وابسته است که شامل توسعه ی یک رویه ی انتقال ژن مؤثر، می باشد. علاوه بر این، استخراج DNA از سلول های سرطانی مناسب و سلول های گیرنده ی مطلوب، برای تشخیص، ضروری می باشد.
در سال 1972، یک رویه ی انتقال مؤثر ژن ابداع شد که بعدها، به آن ترانس فکشن (transfection) گفتند. این رویه که بوسیله ی آن قابلیت ورود مولکول های منفرد DNA به داخل سلول های پستانداران وجود دارد (شکل 1)، بر اساس هم رسوبی DNA با کریستال های کلسیم فسفات، کار می کند. به دلایلی که هم اکنون نیز به خوبی شناخته نشده است، این هم رسوبی می تواند بر روی مقاومت ذاتی سلول ها در برابر مواد ژنتیکی خارجی، فایق آید. این مشخص شده است که سلول های موجود در خط سلول NIH 3T3 که در اصل از فیبروبلاست جنین موش بدست آمده اند، به عنوان DNA خارجی وارد شده به این بخش ها، در نظر گرفته شده است.
این موضوع آخر پیش از این مسئله حل و فصل شد که برنامه ی تجربی بتواند سلول های سرطانی دهنده ای را تشخیص دهد که از آن DNA آماده سازی می شود. در اینجا، محققین کورکورانه عمل کرده اند. این واضح نیست که آیا تمام انواع سلول های سرطانی دارای ژن های تغییر شکل یافته مانند آنکوژن src هستند یا نه؟ همچنین این فهمیده نشده است که آیا یک ژن تغییر شکل یافته ی سلولی (یک آنکوژن سلولی) که تبدیل سلول های اپیتلیال طبیعی به سلول های سرطان است، همچنین قادر به این است که عملکردی غیر مشابه در محیط های بین سلولی مربوط به بافت اتصالی داشته باشد یا نه؟ البته هنوز هم مسائل ممکنه ی دیگری وجود دارد. برای مثال، یک آنکوژن که مسئول تغییر شکل سلول های انسانی نرمال به سلول های سرطانی است ممکن است نتواند سلول های نرمال موش را تغییر شکل دهد، علت این مسئله برخی از ناسازگاری های خاص است.
با در نظر گرفتن این مسائل در ذهن، محققین سلول های توموری دهنده ای را در نظر گرفته اند که از فیبروبلاست موش مشتق شده اند. این سلول های سرطانی خاص از فیبروبلاست موش و از خط سلولی C3H10T1/2 موش مشتق شده اند که به صورت تکراری با استفاده از عوامل سرطان زا و تری متیل کلانترن (3-MC) عمل آوری شده اند. تری متیل کلانترن یک ترکیب شناخته شده از قطران ذغال سنگ است. از این مهم تر، این سلول ها از بخش هایی مشتق شده اند که در آن هیچ عفونت ویروسی و یا ژنوم ویروسی درون زا مشاهده نشده است. در اینجا، هر آنکوژن تغییر شکل دهنده ی تشخیص داده شده در ژنوم این سلول ها، منشأ داخلی دارند. این منشأ داخلی، در واقع نسخه جهش یافته از ژن سلولی نرمال هستند.
در بین سال های 1978- 1979، DNA های مشتق شده از چند خط سلولی تغییر شکل یافته بوسیله ی 3-MC، به داخل محیط کشت های محتوی سلول های گیرنده ی NIH 3T3 انتقال یافت. این مسئله منجر به تولید یک تعداد قابل توجه از کانون های تمرکز سلولی در طی چند هفته می شود. سلول های بدست آمده از بخش متمرکز بعدها موجب ایجاد تومور می شوند (جدول 1). این آزمایش ساده، اثبات کرد که DNA تومور دهنده که حامل یک یا چند المان ژنتیکی است، قادر است تا به صورت سلول گیرنده ی NIH 3T3 غیر توموری تبدیل شود که این سلول ها، به شدت تومورزا هستند.
")
در ابتدا، این به نظر می رسد که تعیین این مسئله سخت است که آیا سلول های توموری دهنده، حامل یک آنکوژن منفرد در ژنوم خود هستند یا چندین آنکوژن متمایز وجود دارد که به صورت هماهنگ عمل کرده و موجب تغییر شمل در سلول های گیرنده می شوند. آنالیز دقیق انجام شده بر روی ترانس فکشن این مسئله را حل کرد. محققین کشف کردند که وقتی DNA سلولی به سلول گیرنده وارد شود، تنها حدود 0.1 % از ارزش ژنوم سلولی مربوط به DNA ی دهنده، وارد ژنوم هر یک از سلول های گیرنده ی تغییر شکل یافته می شود. احتمال اینکه دو ژن دهنده ی غیر مشابه و مستقل از لحاظ ژنتیکی، وارد یک سلول گیرنده ی منفرد شوند، برابر با 〖10〗^3×〖10〗^3 می شود. این احتمال بسیار بعید است. با استفاده از این محاسبات، آنها می توانند استدلال کنند که تنها یک ژن منفرد مسئول تغییر شکل سلول های NIH 3T3 (بعد از ترانس فکشن DNA سلول توموری دهنده) است. در حقیقت، این مسئله منجر این استدلال می شود که سال ها پیش از اینکه برخورد سلول های نرمال C3H10T1/2 موش با عامل سرطان زای 3-MC، موجب تشکیل آلل آنکوژن جهش زای منفرد شود، این آلل ها قادر به تغییر شکل در هر دو سلول C3H10T1/2 و سلول های گیرنده ی NIH 3T3می شود که در این حالت، این آلل ها بوسیله ی انتقال دهنده های ژنی ایجاد می شوند.
این تجربیات مربوط به ترانس فکشن اهمیت بالایی داشته است و بوسیله ی آن، شاخصی مناسب برای مسئله ی آنکوژن ها، مطرح شده است. همچنین این فهمیده شده است که هیچ ارتباط ظاهری میان مکانیزم های مربوط به ژنوم سلولی و عفونت ویروسی وجود ندارد. شاید سلول های تومور انسانی که احتمالا از طریق مکانیزم های غیر ویروسی ظاهری می شوند، همچنین آنکوژن های با قابلیت ترانس فکشن، ایجاد می کنند. آیا آنکوژن های حیوانی (در صورتی که در ژنوم این سلول ها وجود داشته باشد)، همچنین قادر به تغییر رفتار سلول های موش هستند یا نه؟
این سوال و سوالات مشابه به زودی پاسخ داده شد. جواب این سوال ها و سوالات مشابه، جواب مثبت می باشد. DNA های استخراج شده از خط سلولی سرطان های مثانه، ریه و روده یک انسان و DNA مربوط به لوسمی همگی قادر به تغییر شکل در فیبروبلاست NIH3T3 موش گیرنده، هستند (شکل 2). این بدین معناست که آنکوژن های موجود در خطوط سلولی (با هر طبیعتی که دارند)، قادر به تغییر شکل سلولی هستند.
")
آنکوژن های تشخیص داده شده بوسیله ی ترانس فکشن در ژنوم های مختلف سلول های تومور انسانی ظاهرا از ژن های سلولی نرمال قبلی مشتق شده اند که در آنها عملکرد آنکوژن درست نیست. این به نظر می رسد که به موازات این فرایند که منجر به ایجاد رتروویروس های ایجاد کننده ی تغییر شکل، در طی تشکیل این ویروس ها، ژن های سلولی نرمال یعنی پروتو- آنکوژن ها فعال می شوند، ولو اینکه یک مکانیزم ژنتیکی متفاوت رخ دهد.
این توازی ظاهری منجر به مطرح شدن یک سوال واضح می شود: آیا گروه مشابه از پروتو- آنکوژن های سلولی بواسطه ی تلاش رتروویروس ها و سایر عوامل جهش زای غیر ویروسی، فعال سازی می شود؟ یا آیا آنکوژن های مربوط به رتروویروس ها و آنهایی که بوسیله ی مکانیزم های فعال سازی، ایجاد شده اند، از دو گروه متمایز از پروتو- آنکوژن ها می باشند؟
استفاده از پروب های DNA مختص آنکوژن های رتروویروسی، جواب این پاسخ را ارائه می دهد. با استفاده از روش blot جنوبی (Southern blot procedure)، یک پروب DNA که از آنکوژه های H-ras مشتق شده است، قادر به تشخیص و تشکیل ترکیبی از آنکوژن ها می باشد. این آنکوژن ها در DNA مربوط به سلول لوسمی مثانه ی یک انسان، بدست آمده اند (شکل 3). یک آنکوژن مربوطه، که آنکوژن K-ras نامیده می شود و در ژنوم ویروس تومور بدخیم بافت هم بند وجود دارد، قادر است تا با آنکوژن تشخیص داده شده بوسیله ی ترانس فکشن DNA (از یک خط سلولی سرطان روده ی انسانی) تحت تابش قرار گیرد.
")
")
در سال های بعدی، با استفاده از روش های جدیدی مانند روش هیبریداسیون فلورسانس در جا (in situ hybridization (FISH) برای تعیین میزان تقویت ژنی در این نوع و انواع دیگر سلول ها، استفاده شد. در سال 2010، یک سری روش های دیگر برای بررسی و تأیید اثر آنکوژن ها بر روی تقویت ژنی در سرطان های انسانی، مورد استفاده قرار گرفت (شکل 4C). این کار منجر به رسیدن به لیستی 77 تایی از ژن ها شد که به احتمال زیاد، در ایجاد تومور اثر دارند.
")
")
یک وضعیت کاملا متفاوت، بعد از این مشاهده شده است که پروتو- آنکوژن ها بوسیله ی یک رتروویروس کشف شود. بعد از ورود به ژنوم رتروویروس، میزان این ژن دریافت شده، بوسیله ی یک پروموتر رونویسی ویروسی کنترل می شود. این مسئله موجب افزایش میزان ژن می شود. نسخه برداری این ژن مربوط به ویروس، که اکنون به آن آنکوژن گفته می شود، دیگر پاسخی برای سیگنال های سلولی نیست که قبلا میزان آن را تنظیم می کرد. برای مثال، در مورد c-myc میزان این ژن به طور نرمال، بوسیله ی تغییر در سطح سیگنال های خارج سلولی، کنترل می شود مانند چیزی که بوسیله ی فاکتورهای رشد میتوژنی هدایت می شود. وقتی این مورد در ژنوم ویروس میلوسیتوماتوزیز مرغی (avian myelocytomatosis virus (AMV)) مشاهده شود، میزان آن نسبت به حالت نرمال در این سلول ها، بالاست و این میزان در سطح ثابتی قرار دارد.
اما چطور پروتو- آنکوژن H-ras به آنکوژن مناسب تبدیل شود که بوسیله ی ترانس فکشن DNA ی سرطان مثانه ی انسانی، قابل تشخیص باشد؟ تقویت ژنی نمی تواند فعال سازی آن را تضمین کند زیرا این به نظر می رسد که این آنکوژن در DNA سرطان مثانه به عنوان یک ژن کپی شده ی منفرد، وجود دارند. این پازل در زمانی که این آنکوژن سرطان مثانه ی H-ras بوسیله ی کلونی های مولکولی ایزوله شدند، رشد می کند. این آنکوژن در یک بخش DNA ژنومی متمرکز می شوند که دارای طول 6.6 کیلوباز (kilobases) است. این مسئله جالب توجه است که یک بخش DNA با اندازه ی ایده آل، در DNA های انسانی نرمال، مشاهده می شود. بخش آخر به طور واضح بیان کننده ی پروتو- آنکوژن های H-ras می باشد. این آنکوژن یک ژن نرمال است که از برخی جهش ها رنج می برد و در طی تشکیل سرطان مثانه، به آنکوژن تبدیل شده است.
در حالی که ساختارهای کلی DNA بسیار مشابه است، این دو نوع مختلف از ژن H-ras به شیوه ی متفاوتی عمل می کنند. آنکوژنی که از سلول های سرطان مثانه ی انسانی بدست آمده است، موجب تغییر در سلول های NIH 3T3 می شود، در حالی که پروتو- آنکوژن نرمال این قابلیت را ندارند. این راز به نقشه ی جزئی مربوط به مکان های کلیواژی وابسته است که مربوط به آنزیم های محدودکننده ی مختلف می باشد. این مسئله نشان داده است دو نوع مختلف از ژن ها، دارای ساختار فیزیکی هستند که قابل تشخیص هستند.
هنوز این واضح است که دو نوع مختلف از ژن های H-ras دارای برخی تفاوت های قابل توجه در توالی خود هستند. علت این مسئله، عامل های مختلف آنهاست. تفاوت های موجود در توالی بحرانی آنها در اصل بوسیله ی ترکیب مجدد بخش های پروتو- آنکوژن با سایر بخش های مشتق شده از آنکوژن، محدود می شود (شکل 6). این مسئله موجب می شود تا تفاوت حیاتی میان یک بخش، تنها 350 جفت باشد.
")
کشف این جهش نقطه ای، نقطه ی تعیین کننده ای در تحقیقات سرطان می باشد. این اولین زمان بود که یک جهش در یک ژن کشف شد که موجب رشد نئوپلاستیک سرطان در انسان می شود. این مسئله نیز مهم است که این تغییر ژنتیکی به عنوان یک جهش سوماتیک (somatic mutation) در نظر گرفته می شود.
")
از این مهم تر، این جایگزینی جفتی در چارچوب ژن H-ras رخ می دهد. به طور خاص، این جهش نقطه ای موجب جایگزینی یک باقیمانده ی گلیسین در باقیمانده های والین مربوط به پروتئین های کد گذاری شده ی H-ras می شود.
کشف این جهش نقطه ای، یک مکانیزم در مورد فعالیت انکوژنی ارائه کرده است که کاملا از فعالیت انکوژنهای MYC متفاوت است. در مورد H-ras، یک تغییر در ساختار مربوط به پروتئین کدگذاری شده، یک مورد حیاتی می باشد. برخلاف آنکوژن MYC، تنظیم نبودن میزان این ماده در قدرت
استفاده از مطالب این مقاله، با ذکر منبع راسخون، بلامانع می باشد.










! / استاد توکلی")













